
Contents
- Difference in control processes for unimanual and bimanual movements
- Mechanism for coordinated bimanual movement
- Learning mechanism of rhythmic movements
- Effect of motor plan on motor learning
Research
Difference in control processes for unimanual and bimanual movements
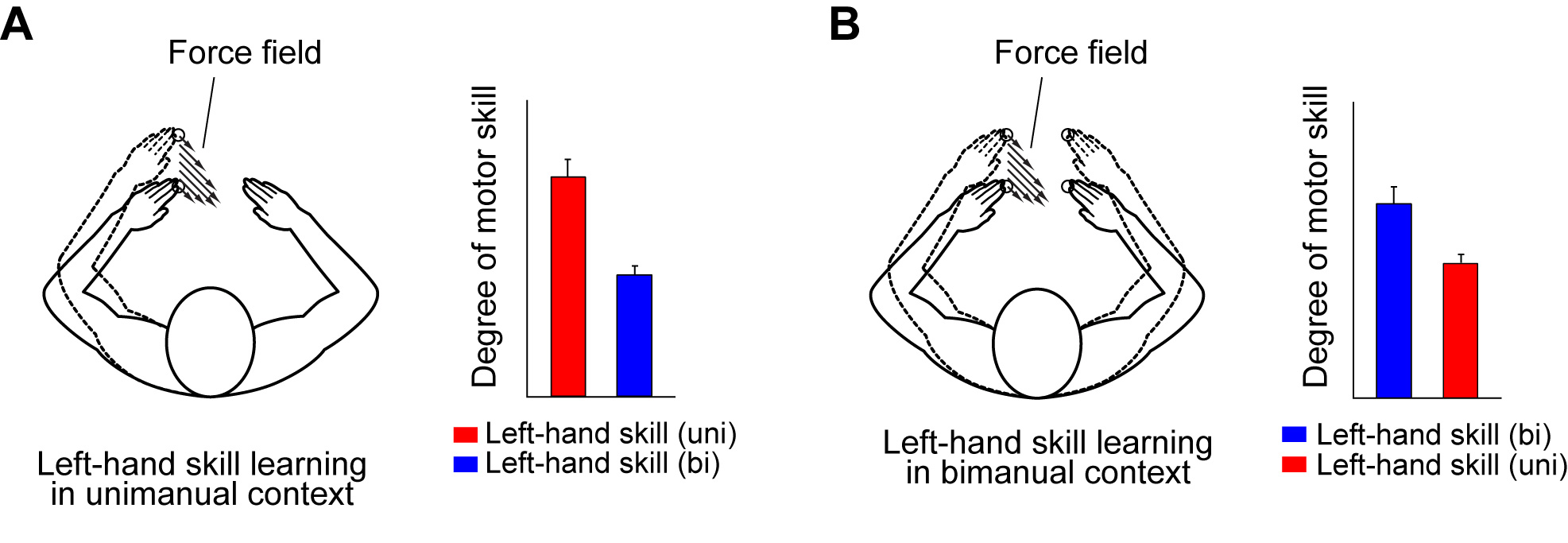
A limb’s motion appears to be similar across unimanual and bimanual movements. However, we demonstrated partial, but not complete, transfer of learning across these behavioral contexts.
After participants were trained to move thier left hand straightforward unimanually in the presence of the force field (Fig. A), we examined whether the learned motor skill in unimanual context is transfered to the same left-hand motion in bimanual context. Although the required motor command for the left hand is the same between unimanual and bimanual contexts, participants could not fully exhibit the learned motor skill in bimanual context (Fig. A). It was also found that the left-hand motor skill learned in bimanual context (blue in Fig. B) is not fully transfered to the same left-hand motion in unimanual context (red in Fig. B). These results suggest the difference in control processes for the same limb motion between unimanual and bimanual contexts.
 Original article: Nozaki, Kurtzer, and Scott (2006) Nature Neuroscience.
Original article: Nozaki, Kurtzer, and Scott (2006) Nature Neuroscience.
Mechanism for coordinated bimanual movement
Coordinated bimanual movement requires the neural controller (internal model) for each arm to exchange information on each other's movement. How does each controller integrate information on both arms' movements?
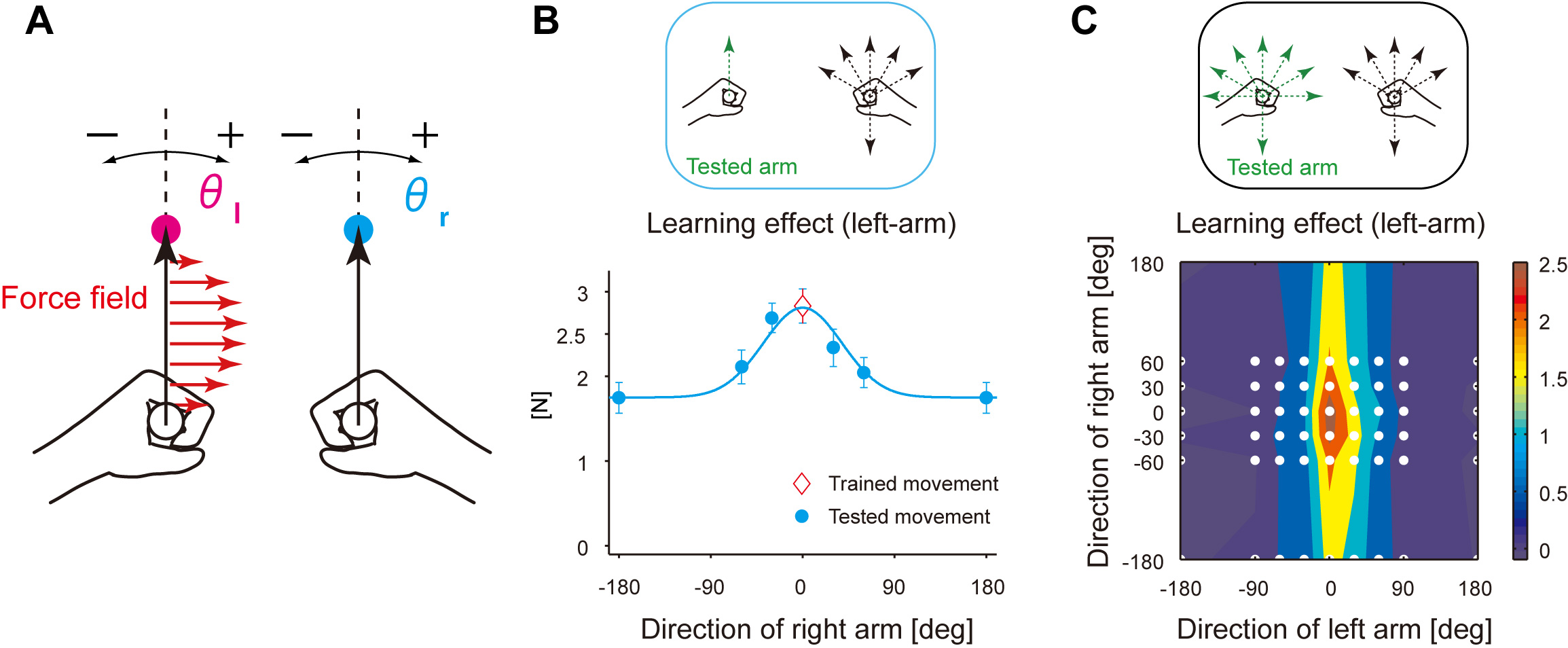
First, we examined how the control of an arm movement in bimanual context is affected by the other arm movement. Human subjects were instructed to move both hands forward simultanesouly with holding the handles of robotic arms (Fig. A). The force field was applied only to the left handle, and subjects were trained until they leanred to move both hands forward in the presence of the force filed (Fig. A). Figure B shows the generalization pattern of the left-hand motor learning observed when changing the movement direction of the right arm, indicating that it gradually decayed as the direction of right arm movement is away from the foward direction (Fig. B).
The generalization pattern observed when changing the movement direction of both arms was consistent with the pattern predicted from the gain-field encoding scheme in which the movement directions of both arms are multiplicatively integrated (Fig. C).
Importantly, the multiplicative encoding scheme theoretically predicts that participants could adapt reaching movements to complicated force fields depending nonlinearly on the movement directions of both arms. Indeed, our participants could readily adapt such complicated force fields, which has been conventionally considered to be impossible in motor learning litereture.These results indicate that multiplicative encoding of the kinematics of both arms enables the brain to control bimanual movement by providing great flexible ability to handle arbitrary dynamical environments resulting from the interactions of both arms.
 Original article: Yokoi, Hirashima, and Nozaki (2011) Journal of Neuroscience.
Original article: Yokoi, Hirashima, and Nozaki (2011) Journal of Neuroscience.
Learning mechanism of rhythmic movements
When performing a discrete movement such as pitching a baseball, it is known that the brain can promote learning by modifying the motor commands of successive movements based on the difference (error) between the intended and actual results. On the other hand, the brain mechanisms in rhythmic, repetitive movements, such as dribbling a basketball, are not well understood. During rhythmic movement the brain receives error information on a continuous basis. Is the brain’s error-correction mechanism able to work as well as it does for discrete movements?
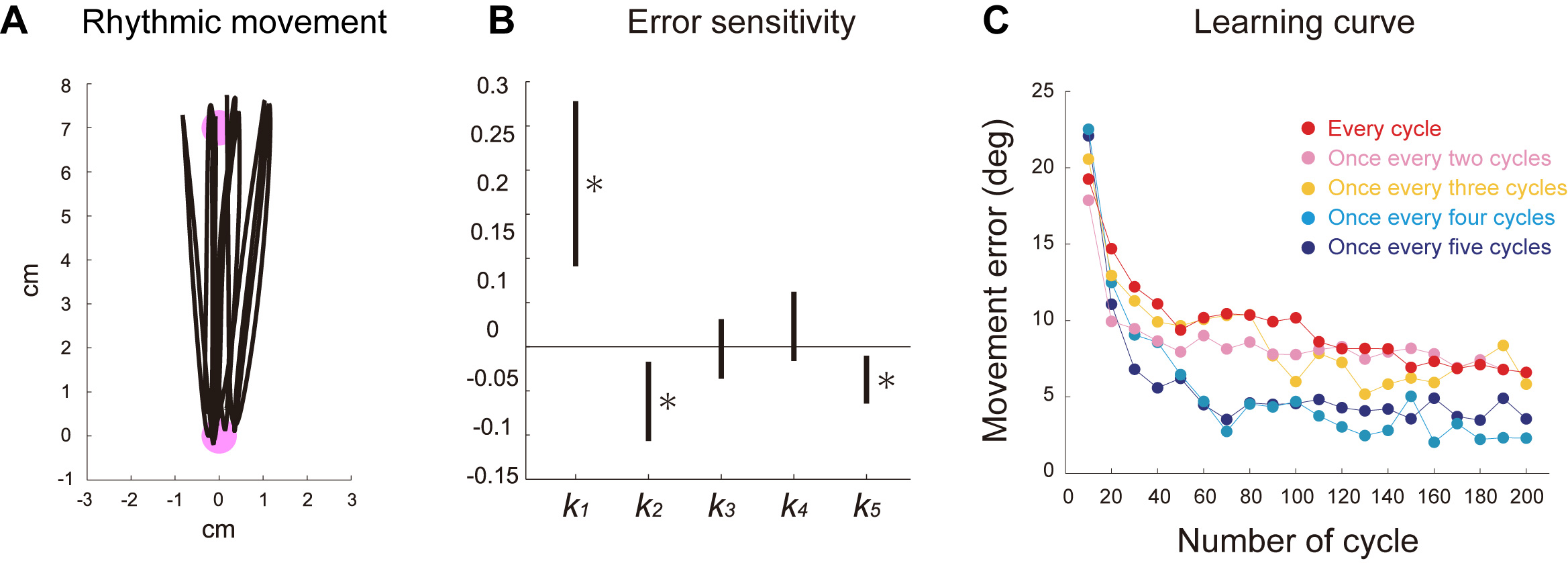
We had human participants perform rhythmic movements between two targets (Fig. A) and examined how the error information of each cycle affects subsequent cycles using system identification technique. We found that error information obtained from one cycle not only appropriately modified the next cycle movement but also deteriorated several later cycles (Fig. B).
This result led to a counterintuitive prediction: providing visual error feedback for only a fraction of cycles should enhance motor learning. As predicted, we observed that motor adaptation to a constant visual rotation (30°) was significantly enhanced by providing visual feedback once every fourth or fifth cycle rather than for every cycle. This result suggests that the brain is not able to deal appropriately with the overwhelming flow of error information generated during rhythmic movements.
 Original article: Ikegami, Hirashima, Osu, and Nozaki (2012) Journal of Neuroscience.
Todai Research:When watching too closely is a bad thing.
-Intermittent visual feedback can improve learning of rhythmic movements-
Original article: Ikegami, Hirashima, Osu, and Nozaki (2012) Journal of Neuroscience.
Todai Research:When watching too closely is a bad thing.
-Intermittent visual feedback can improve learning of rhythmic movements-
Effect of motor plan on motor learning
It is well-established that the brain internally represents the dynamics of the body and environment as an internal model for feedforward control of movements. Although, conventionally, it has been thought that only one internal model can be learned for a movement kinematics (e.g., straightforward reaching movement), we demonstrated that the brain can learn distinct internal models for identical movements according to distinct motor plans.
First, we created a seemingly impossible situation where human participants perform physically identical movement irrespective of thier intention (or motor plans). While participants reached to each of two targets located at a clockwise or counter-clockwise position, a gradually increasing visual rotation was applied in the clockwise or counter-clockwise direction, respectively, to the onscreen cursor representing the unseen hand position. As the visuomotor (VM) rotation angle increased, participants gradually adapted to the VM rotations; the actual hand trajectory gradually approached the straightforward direction (right in Fig. A), whereas the cursor trajectory on the screen was kept almost constant (left in Fig. A). Thus, this procedure could implicitly led participants to perform physically identical reaching movements irrespective of their intentions (i.e., movement plans) to move their hand toward two distinct visual targets.
To examine whether these physically identical movements could learn distinct internal models, we applied CW and CCW force fields for movements toward the CW and CCW targets, respectively (Fig. B). If only one internal model is allowed to be leanred for physically identical movement, participants could not adapt to the opposing force fields. However, contrary to the conventional notion, our participants could readily adapt to them (Fig. C). This result clearly indicates that multiple internal models can be learned and flexibly retrieved, even for physically identical movements, according to distinct motor plans in a visual space.
![]() Original article: Hirashima and Nozaki (2012) Current Biology.
Original article: Hirashima and Nozaki (2012) Current Biology.