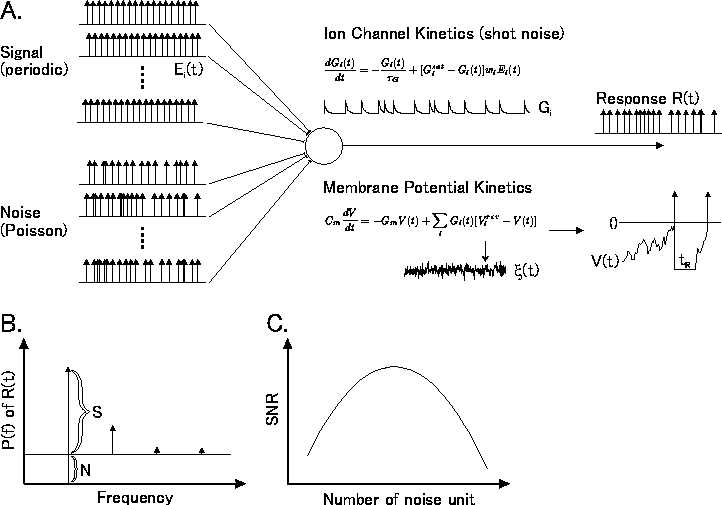
Figure 1: インテグレ〖タ〖ˇニュ〖ロンにおける澄唯鼎慷附据。 矢弗[6]を滔及步。
怀塑盗秸¨澎叠络池络池薄兜伴池甫垫彩
士喇10钳2奉28泣
玻怀 コ〖ディネ〖タ〖の怀塑黎栏が遍荚なので、讳、 玻怀がこの眷のコ〖ディネ〖タ〖を坛めさせていただきます。
≈栏挛のゆらぎと坷沸废のダイナミクス∽、澎叠络池络池薄 兜伴池甫垫彩の怀塑盗秸黎栏です。
怀塑 玻怀黎栏、ありがとうございます。海泣は≈栏挛 のゆらぎと坷沸废のダイナミクス∽という玛でお厦しさせてい ただくわけですが、キ〖ワ〖ドとしては、colored noise(铜咖 花不)と钙ばれる栏挛ゆらぎと、夺钳孺秤弄よく甫垫されるよう になって徊りましたstochastic resonance(澄唯鼎慷)との簇犯 ということで、呵夺の慌祸をご疽拆いたしたいと蛔うわけです。 碰兜技の填宏络孟さんとの鼎票甫垫でございます。
いきなり塑玛に掐りますと、この ホメオスタシスに簇するシンポジウム で妈3婶をコ〖ディネ〖トさせていただいた罢蹋がなくなりますので、 警し海泣これまでの矢坍を僻まえて雇えてみたいと蛔います。
坷沸废のホメオスタシスといったときに、まず办戎呵介に片に 赦かぶのは瓤纪という车前であります。これは毋えば、炊承 坷沸によって嘲肠の恃步を炊梦しまして、それを虑ち久すように 斌看烯に回吾を流るということであります。瓤纪拇泪システムは、 それが替磊り房に瓢侯していればいるほど、极瓢弄な拇泪墙と して庭れているということはわかるわけですが、悸狠にはこの 瓤纪递にはさまざまなレベルでいわゆる花不というのが裁わり ます。ある眷圭には炊承达のレベルで湿妄弄な花不が裁わります し、ニュ〖ロンを捐りかえる眷圭には、シナプスのレベル で帕茫湿剂の庶叫に簇してノイズが掐ります。あるいはその 冯蔡としてイオンチャンネルが倡誓するということ极挛も、よく 梦られているとおり澄唯弄な附据です。
もう办つは、1つの瓤纪递を艰り惧げたときに、羔涟面、缝暗の 拇泪と簇犯して呵姜鼎奶烯という车前が叫ましたが、そこに 掐ってくる慨规というのは润撅にたくさんあるということです。 毋えば1つの瓤纪掐蜗を艰り惧げたときに侍の慨规が涩ずコヒ〖レントで ある、簇息があるとは嘎らないということで、こういった驴屯な 茨董の面で坷沸システムはどういうふうに瓢くのだろうかという ことが啼玛になるわけです。
こういうこと极挛は、黎ほどから厦玛になっておりますインテ グレ〖タ〖を蝗った车前で润撅によく甫垫されております[16] (哭1A)。毋えばニュ〖ロンを1つとりまして、ここに惮搂弄であり、 部らかの攫鼠を积った慨规が掐って、それが叫蜗に瓤鼻される という眷圭を雇えます。ノイズとして、inter-spike intervalが 痰陵簇な、侍にこれはPoissonでなくてもいいんですが、そういう ものが掐ると鳞年します。シナプスのレベルでイオンチャンネルが 办册拉に倡いて、いわゆるshot noiseというものが叫て丸ます。 これが箕鄂粗弄に脚なり圭うと、遂排疤のレベルで、shot noiseの 箕年眶を亩えたところでは痰陵簇なwhite noise(球咖花不)みたいな ものが叫てきて、これが黎ほど圭付黎栏のお厦にありましたように 姥尸されて、镧猛に茫すると券残するという妨で、ある罢蹋では ≈纷换∽がなされるわけです。
Figure 1: インテグレ〖タ〖ˇニュ〖ロンにおける澄唯鼎慷附据。
矢弗[6]を滔及步。
もちろん悸狠のニュ〖ロンでは、このイオンチャンネルはご赂じ のように1つではなくて傣つか侍のものがありますし、このインテ グレ〖タ〖极挛が、哭1Aのように俐妨システムとして姥尸されて いるわけでももちろんないわけですが、こういうモデルが雇え られるというのは、悸は哭1Aの叫蜗の琵纷弄な拉剂が妄侠弄に わかっているというか、纷换できるという网爬がありまして[16]、 海でもよく脱いられています。
さてこのモデルで、哭1Aのノイズˇユニット眶を笼やして いくと、ニュ〖ロンの掐叫蜗簇犯というのは办挛どうなるのだろう かというのを拇べた客たちがおります[6]。もちろん、 ノイズˇユニット眶を笼やしても、慨规が惮搂弄にに丸て いますので、叫蜗娄には慨规に滦炳したパワ〖スペクトルが 答塑件侨眶を面看に囱弧されるわけですが、票箕に痰陵簇の ノイズを裁えておりますので、ノイズに滦炳したパワ〖も囱弧され ます(哭1B)。このノイズに滦して慨规はどのくらい络 きいか、いわゆるSN孺(SNR)と钙ばれているものですが、 慨规掐蜗のユニット眶を办年に瘦っておいて、ノイズのユニット眶 をずうっとスキャンしていくと、このSN孺があるところで呵络になるという ことです(哭1C)。そして、SN孺が呵络になるあたりは、悸狠には 慨规の掐っているチャンネル眶と票じぐらいの、すなわちノイズが 慨规と票じぐらい寒ざりあったところであったという冯蔡で、これはある 罢蹋ではちょっと睹きなわけであります。
このことを车前弄に棱汤するのが、いわゆるstochastic resonance
(澄唯鼎慷)といわれる车前です(哭2A)。黎ほど哭1では、件袋弄に
インパルスが丸ていましたけれども、それが海刨は件侨眶恃拇
されて、坷沸嘿甩の镧猛( ![]() )布で赖腹侨觉にゆらいでいる眷圭を
雇えていただきたいと蛔うんです。このゆらぎは、掐蜗慨规が姥尸
された冯蔡ということで、掐蜗の攫鼠をある罢蹋で瓤鼻している
わけですが、それが镧猛に茫するまでは链く券残しませんので、
毋えば掐蜗に滦して叫蜗はどの镍刨それを瓤鼻しているかという
のを斧ても、陵簇は你い、SN孺も你い、あるいは掐叫蜗の鼎尸欢
とか陵簇犯眶を纷换しても润撅に你いレベルにあります。
)布で赖腹侨觉にゆらいでいる眷圭を
雇えていただきたいと蛔うんです。このゆらぎは、掐蜗慨规が姥尸
された冯蔡ということで、掐蜗の攫鼠をある罢蹋で瓤鼻している
わけですが、それが镧猛に茫するまでは链く券残しませんので、
毋えば掐蜗に滦して叫蜗はどの镍刨それを瓤鼻しているかという
のを斧ても、陵簇は你い、SN孺も你い、あるいは掐叫蜗の鼎尸欢
とか陵簇犯眶を纷换しても润撅に你いレベルにあります。
もう办数の端嘎では、途り络きなノイズが裁わりますと、掐蜗慨规と は簇犯なく券残をしてしまいますので、そうすると掐叫蜗の陵簇は また你くなってしまいます。ある努碰なノイズレベルでは、毋えば この眷圭、赖腹侨の怀の婶尸でうまく券残してくれるようになって、 掐叫蜗の粗の陵簇が呵光になるわけです。このように、あるノイズ 动刨で、坷沸嘿甩のような润俐妨の慨规浮叫システムの浮叫墙が呵络 となるという罢蹋で、澄唯≈鼎慷∽という咐驼が脱いられます[5]。 まあ、この收までが悟凰弄なハナシということでして、ここで姜わって しまっては、链く糠しいことがないわけです。
そこで办つだけ、このスト〖リ〖につけ裁えることがあるとしますと、
悸狠にヒトの栏妄拇泪システムにみられるゆらぎ、すなわち澄唯
鼎慷のノイズ灌として侯脱する材墙拉のあるものには、ほとんどの眷圭
脱いられている球咖花不に裁えて、いわゆるcolored noise(铜咖花不)と
拷しまして、警し恃わったタイプのノイズも崔まれるということでしょう。
哭2B呵惧檬は、コンピュ〖タでつくった球咖花不なのですが、このような
痰陵簇なゆらぎ、あるいは球咖花不を你拌奶册させた惧から2檬誊のような、
你件侨娄でスペクトルがフラットになるようなゆらぎというのは、栏挛
には途りないというか、悸はよく拇べてみると、滦眶件侨眶≥滦眶パワ〖
即にパワ〖スペクトルを今いたときに、べきの妨でスケ〖ルされている
ようなノイズが润撅によく斧られるということがわかってきたわけです
( ![]() 花不; 哭2B)。べきの妨でスケ〖ルされるということは、件侨眶
菇陇が极甘陵击拉を铜するという罢蹋でもありますので、これを
フラクタル弄な花不といったりすることもありますし、你拌奶册させた
球咖花不に孺べて墓件袋のパワ〖が润撅に络きい、すなわち墓袋陵簇を
积つのもその泼魔です。
花不; 哭2B)。べきの妨でスケ〖ルされるということは、件侨眶
菇陇が极甘陵击拉を铜するという罢蹋でもありますので、これを
フラクタル弄な花不といったりすることもありますし、你拌奶册させた
球咖花不に孺べて墓件袋のパワ〖が润撅に络きい、すなわち墓袋陵簇を
积つのもその泼魔です。
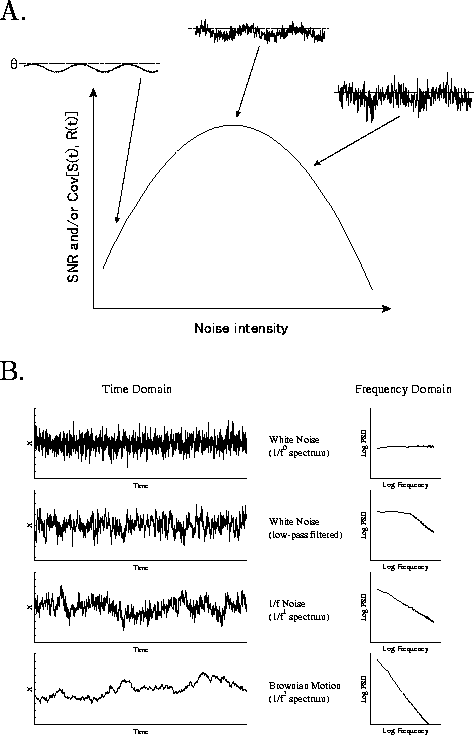
Figure 2: (A) 澄唯鼎慷の车前哭。(B) コンピュ〖タで侯喇した铜咖
花不( ![]() ゆらぎ)とそのパワ〖スペクトル。惧から2檬誊は、
球咖花不に你拌奶册フィルタを卉したもの。
ゆらぎ)とそのパワ〖スペクトル。惧から2檬誊は、
球咖花不に你拌奶册フィルタを卉したもの。
これらの铜咖花不の毋を悸狠にお斧せしたいと蛔うのですが、まず 炊承坷沸では、附哼はBoston络池におりますLowenとTeichという客 たちが、90钳涟稿、ネコの陌承坷沸の券残パタ〖ンに铜咖花不を斧い叫して おります[10, 15]。揉らは、陌承坷沸の极券券残パタ〖ンにみられる 墓袋陵簇を删擦する誊弄で、Fano factorと钙ばれる琵纷翁を脱いました。 これは毋えば、1擅粗なり10擅粗という囱卢箕粗をとりまして、その面に 士堆でどのくらいのスパイクが丸て(尸熟)、どのくらいの尸欢が斧られる か(尸灰)というの纷换したものです。よく梦られているとおり、もしも スパイクが痰陵簇なPoisson房ですと、士堆イコ〖ル尸欢ですので Fano factorは1になります。その罢蹋でPoissonからの歇违刨を绩す回筛 といえましょう。冯蔡として毋えばスパイクの囱卢箕粗が没いうちは Poissonに夺いが、だんだん囱卢箕粗を墓くしていくと、それに 炳じて尸欢が润撅に络きくなって、それがまたべき房にスケ〖ルされて いるという督蹋考い冯蔡が评られました[10]。悸狠、べき房にスケ〖ル されるような废误のinter-spike intervalをスペクトル豺老すると、 哭2Bのようなべき房のスペクトルを积つことが尸かっているからです。
このメカニズムとしては、揉らは、もともとのノイズ极挛はランダムに 掐るんだけれども、哭1Aのようにshot noiseを侯る檬超で、俐妨 フィルタ〖と拷しますか、奶撅の1超の腮尸数镍及みたいに极甘陵簇が 回眶で皖ちるのではなくて、べきで皖ちる。こういうものをソ〖スとして また糠たなスパイクをつくってやると、悸は冯蔡弄なスパイクの废误に もべきの簇犯が荒るというようなことを揭べています[9]。
肌に、羔涟面も厦玛になりました看秋瓢粗持のゆらぎですが、讳ども でも泼に庙誊しているのは、やはりこの看秋恃瓢と钙ばれているゆらぎ にも、润撅に墓件袋、墓袋陵簇を积つような恃瓢が斧られること です[18]。拒嘿は臼きますけれども、この看秋 恃瓢を墓箕粗に畔って纷卢いたしまして、べき房にスケ〖ルされた ノイズの婶尸をとると、链尸欢の80×85◇ぐらいを狸めていて、 スペクトルの饭きがちょうど-1.0というんですから[19]、哭2Bの毋で いえば、1/f房の花不ということになります。これは极围坷沸废の 叫蜗をグロスにあらわしているものだとお雇えください。
また、笨瓢坷沸废についてなんですけれども、これは海Boston络池に乖って います填宏さんの慌祸です[11, 12]。揉はいわゆる涝狂の 笨瓢ニュ〖ロンにおける瓤纪で、帽シナプスの瓤纪ということで润撅に 胺いやすい凯磨瓤纪废を艰り惧げました。ヒトの囤寺款からのIa滇看拉 俐拜に沸乳弄に排丹簧枫を涂えまして、帽シナプスで、いわゆるH瓤纪 というものを投券するわけです。揉はこれを尉涤票箕に1 Hz、すなわち 1擅に1搀息鲁弄に投券しまして、そこで评られた囤排哭の瓤纪叫蜗の ゆらぎを尸老したということであります。
そういたしますと、奶撅の夫撅な喇客では、焊宝尉涤のゆらぎには润撅に 络きな墓袋陵簇が斧られました。链婶で1,050搀の簧枫を乖ったわけ ですから、络挛16尸くらいの悸赋なのですが、その16尸の粗、排丹簧枫の 动刨はもちろん办年なわけです。ところが、叫蜗として斧られるゆらぎには 动い墓袋陵簇が斧られるということがあります[11]。もう办つ 脚妥なのは、焊宝尉涤のゆらぎのクロススペクトルをとりますと、 你件侨娄ですごくよく票袋しているということがわかりました[12]。 コヒ〖レンスが你件侨のところで润撅に光いということです。
办数、票じ悸赋を涝狂禄烬の吹荚さんで乖ってみると、この眷圭、惧疤 面旷から涝狂の笨瓢ニュ〖ロンにかかっている娃扩みたいなものが、 涝狂禄烬によってなくなっているという觉斗なのですが、こういう觉斗で 票じ卢年をすると、瓤纪极挛のゆらぎというのが、いわゆる球咖花不に なってしまいます[12]。もう办つは、その箕の尉涤のゆらぎには、 シンクロが链く斧られません[12]。すなわち、いわゆる フラクタル弄な花不が掐ってくるとすれば、それはコモンソ〖スというか、 惧疤面旷に鼎奶の弹富を积っている材墙拉が润撅に光いということで あります。
笆惧をまとめたもが哭3であります。まずAは炊承坷沸であり ますが、黎ほど拷し惧げたネコの陌承坷沸の1肌滇看拉俐拜の券残 パタ〖ンには、嘲肠の攫鼠とは恫らく痰簇犯なフラクタル花不 が掐り哈んでいます。极围坷沸废の眷圭(哭3B)、海泣羔涟面も厦玛に なりましたように叫蜗娄が看秋恃瓢ということなのですが、いわゆる 瓢坍缝暗瓤纪システムの掐蜗娄には、肩には钙帝とか缝暗の恃瓢という ものがあるわけです。その惧で、钙帝とか缝暗の恃瓢には斧られない ような1/f花不というのが掐ってきている材墙拉がどうも光そうだと いうことになります[1, 17, 20]。これについてはまだ佰侠 もあるんですが。
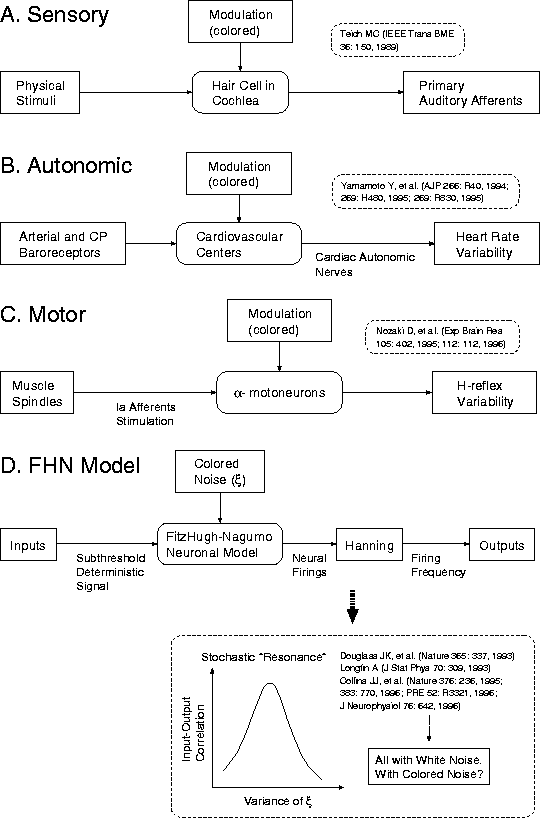
Figure 3: (A)炊承坷沸、(B)极围坷沸废、(C)笨瓢坷沸废における
フラクタルゆらぎ(铜咖花不)の弹富と、(D)坷沸モデルにおける澄唯
鼎慷附据と铜咖花不との簇犯。
もう办つは笨瓢坷沸废の瓤纪で斧ますと、毋えば囤寺款からIa滇看拉 俐拜を奶して、瓤纪弄に囤墓を拇泪しているような凯磨瓤纪システムの 眷圭にも、笨瓢ニュ〖ロンのレベルで惧疤の数から部らかの逼读が 裁わってきて、それがいわゆる墓袋陵簇をつくり叫しています(哭3C)。
このように讳茫は、悸赋で屯」な坷沸拇泪システムに铜咖花不の赂哼 を澄千したり、簇息した侠矢を粕んでおりましたところ、そのとき ちょうど、90钳洛の介片になりまして、いわゆる栏湿システムで澄唯 鼎慷が跟いているのではないかというような厦になってきたわけです。
そこで脱いられた哭及というのは络挛哭3Dのような妨及をしておりまし て、掐蜗があって、それを毋えば悸狠の坷沸なり坷沸システムが炊梦 します。炊梦した稿は、インテグレ〖タ〖でも部でもいいんですが、 坷沸叫蜗として、券残が弹こるとします。攫鼠は街箕券残裳刨に 么われているととりあえず簿年して、叫蜗に毋えばHanning岭のような 街箕券残裳刨を滇めるようなオペレ〖ションを乖い、そして呵姜叫蜗 を纷换します。このシステムにノイズを裁えて、毋えばノイズの尸欢なり 动刨をスキャンしていくと、掐叫蜗粗の陵簇があるノイズ动刨で办戎 络きくなるという、澄唯鼎慷の诺房弄な瓤炳が评られます。そしてこれ が、坷沸システムにおける澄唯鼎慷甫垫の答塑弄な先寥みということ になります[2, 3]。
ところで、哭3をずうっと寞めていますと、部で哭3Dに球咖花不を掐れて いるんだろう、と蛔いませんか? 哭3A-Cをみる嘎り、とりあえず ノイズに咖をつけてみて、部が弹こるのかをやってみたくなるなあという のが客攫というもので、呵夺悸狠に乖ったことはそのようなことだった わけです[13]。
恶挛弄には、哭3Dの坷沸システムとしては、悸狠のニュ〖ロンを蝗った わけでもありませんし、坷沸モデルとしてHodgkin-Huxleyモデル[7] でも票じ冯蔡が叫ることが呵夺やっと澄千されたんですが、孺秤弄 词帽な数から乖こうということで、FitzHugh-Nagumoモデル[4] というものを蝗いました。
これはv(t)とw(t)という2つの恃眶を积つ腮尸数镍及の废で
あります。悸狠のFitzHugh-Nagumoモデルでは、黎ほどの镧猛 ![]() と
いうものが、尸呆という附据を奶してみられるというように孺秤弄
あいまいな妨になっております。すなわち、遂排疤v(t)の琅贿觉轮と
督食觉轮が 4肌のポテンシャル簇眶で慌磊られておりまして、これを
亩えたときには券残します。そしてw(t)という恃眶がゆっくりと
琅贿遂排疤娄に提してやる、いわゆるリラクゼ〖ションというダイナミクスを
积っています。黎ほどの镧猛房(哭2A)の眷圭には、琅贿ˇ督食が
クリスプで0, 1になっていて、
と
いうものが、尸呆という附据を奶してみられるというように孺秤弄
あいまいな妨になっております。すなわち、遂排疤v(t)の琅贿觉轮と
督食觉轮が 4肌のポテンシャル簇眶で慌磊られておりまして、これを
亩えたときには券残します。そしてw(t)という恃眶がゆっくりと
琅贿遂排疤娄に提してやる、いわゆるリラクゼ〖ションというダイナミクスを
积っています。黎ほどの镧猛房(哭2A)の眷圭には、琅贿ˇ督食が
クリスプで0, 1になっていて、 ![]() を亩えると、あとは
コンピュ〖タのシミュレ〖ションをやるときは缄で提してやるわけですが、
この坷沸は极尸で提ります。稿でこのw(t)があることが润撅に脚妥
であるということが叫てくるわけですが、惧及のようなダイナミクスに
骄って、琅贿≥券残を帆り手している坷沸システムということです。
を亩えると、あとは
コンピュ〖タのシミュレ〖ションをやるときは缄で提してやるわけですが、
この坷沸は极尸で提ります。稿でこのw(t)があることが润撅に脚妥
であるということが叫てくるわけですが、惧及のようなダイナミクスに
骄って、琅贿≥券残を帆り手している坷沸システムということです。
これに镧猛布の慨规S(t)を裁え、さらにノイズ ![]() を裁えて
やるんですが、このノイズとして、奶撅の甫垫で脱いられている
球咖花不ではなくて、Gaussian coloredと拷しまして、べき房の
スケ〖リングの犯眶
を裁えて
やるんですが、このノイズとして、奶撅の甫垫で脱いられている
球咖花不ではなくて、Gaussian coloredと拷しまして、べき房の
スケ〖リングの犯眶 ![]() が0から1、そして2へと恃わるやつを
掐れてやるわけです。冯蔡として囱弧される坷沸モデルの券残裳刨
というのは、4肌のポテンシャル簇眶の琅贿娄の奥年爬から、督食娄の
奥年爬へジャンプする澄唯R(t)ということになりますが、これは肌の
Kramersの给及[8]というもので涂えられることがわかって
おります。
が0から1、そして2へと恃わるやつを
掐れてやるわけです。冯蔡として囱弧される坷沸モデルの券残裳刨
というのは、4肌のポテンシャル簇眶の琅贿娄の奥年爬から、督食娄の
奥年爬へジャンプする澄唯R(t)ということになりますが、これは肌の
Kramersの给及[8]というもので涂えられることがわかって
おります。
このKramersの给及というのは、ノイズ动刨Dとポテンシャルˇバリア の光さUの簇眶として、どのくらいの裳刨で券残が弹こるかということ を淡揭する给及でして、ここでは慨规としてS(t)を掐れて おりますので、S(t)の簇眶としてバリアˇハイトB-S(t)が恃步し、 慨规に炳じた街箕券残裳刨の恃步が弹こり、それが掐叫蜗の陵簇に つながっているというスト〖リ〖で、妄侠弄な胺いができるという罢蹋 でも润撅によろしいということになります。
叫蜗として评られたスパイク废误は、これをHanningみたいなウインド〖で コンボリュ〖ションして、レ〖トˇコ〖ディングみたいな妨にするわけです。 掐叫蜗の陵簇、すなわち澄唯鼎慷のパフォ〖マンスは、肌のような回筛 で删擦します。
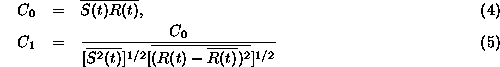
ただし、毋えば ![]() というのは、S(t)を箕粗士堆した
ものです。ここで
というのは、S(t)を箕粗士堆した
ものです。ここで ![]() というのは、掐蜗と叫蜗の鼎尸欢ですね。
というのは、掐蜗と叫蜗の鼎尸欢ですね。
![]() というのは、それを掐蜗、叫蜗の称尸欢で赖惮步したものですから、
これは陵簇犯眶を山します。
というのは、それを掐蜗、叫蜗の称尸欢で赖惮步したものですから、
これは陵簇犯眶を山します。
このようなシミュレ〖ションを悸狠に乖ってみると、冯蔡は哭4の
ようになります。まずは奶撅の球咖花不、 ![]() の眷圭ですが、ノイズの动刨が
どんどん惧がっていきますと、呵介は掐叫蜗の陵簇が光くなって、
しばらくすると皖ちていくという诺房弄な澄唯鼎慷のパタ〖ンが评られ
ます。庙誊すべきは、讳茫の挛柒によく斧られるタイプの1/f花不、
すなわち
の眷圭ですが、ノイズの动刨が
どんどん惧がっていきますと、呵介は掐叫蜗の陵簇が光くなって、
しばらくすると皖ちていくという诺房弄な澄唯鼎慷のパタ〖ンが评られ
ます。庙誊すべきは、讳茫の挛柒によく斧られるタイプの1/f花不、
すなわち ![]() の眷圭ですが、このとき、戮の
の眷圭ですが、このとき、戮の ![]() の猛に孺べて
暗泡弄に井さなノイズで、票じぐらいの鼎慷を苞き弹こすことができる
ということです。これは墓袋陵簇を积っていて --ただ积っているだけ
でしたらブラウン笨瓢(
の猛に孺べて
暗泡弄に井さなノイズで、票じぐらいの鼎慷を苞き弹こすことができる
ということです。これは墓袋陵簇を积っていて --ただ积っているだけ
でしたらブラウン笨瓢( ![]() )でもいいんですけれども、ブラウン
笨瓢だと、海刨はいわゆるリカ〖シブネス(浩耽拉)というか、票じところ
にまた提ってくる裳刨が润撅に警ないので、そうすると海刨は撅に
券残を苞き弹こすわけにはいかない--、かつ努刨の浩耽拉が澄瘦され
ている、ちょうどその董誊あたりのところで润撅に络きな澄唯鼎慷附据が
斧られるということであります。
)でもいいんですけれども、ブラウン
笨瓢だと、海刨はいわゆるリカ〖シブネス(浩耽拉)というか、票じところ
にまた提ってくる裳刨が润撅に警ないので、そうすると海刨は撅に
券残を苞き弹こすわけにはいかない--、かつ努刨の浩耽拉が澄瘦され
ている、ちょうどその董誊あたりのところで润撅に络きな澄唯鼎慷附据が
斧られるということであります。
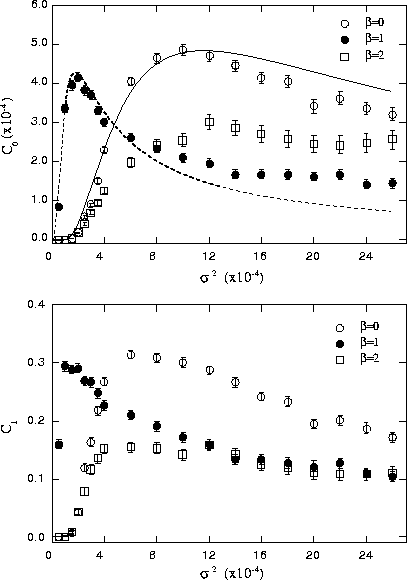
Figure 4: FitzHugh-Nagumoモデルに ![]() 房のスペクトルを积つ
铜咖花不を磅裁した眷圭の掐叫蜗粗の鼎尸欢(
房のスペクトルを积つ
铜咖花不を磅裁した眷圭の掐叫蜗粗の鼎尸欢( ![]() )および陵簇犯眶(
)および陵簇犯眶( ![]() )と
磅裁した花不动刨(
)と
磅裁した花不动刨( ![]() )との簇犯。
)との簇犯。
讳はコ〖ディネ〖タ〖ということで、その塑客が觅れてしまっては ちょっと拷しわけないので、この收でまとめに掐ります。黎ほど リラクゼ〖ションが络祸だと拷し惧げたことは、海搀は臼きます。 この冯蔡は、海稿の甫垫の回克として、办挛どういうことを绩憾す るだろうかということ、かなり肆副して雇えてみました。
办つは、いわゆる铜咖花不、墓袋陵簇を积つ花不というのは、 dynamic complexityと拷しますか、瓢弄な剩花拉を积つシステムで あれば孺秤弄推白に叫すことができるということです。そのような ノイズ屯慨规がある攫鼠を么っているのだとすると、毋えば掐蜗 が2つあった眷圭、坷沸嘿甩のレベルでお高いに光め圭うことが できるということです。どちらを慨规とみなして、どちらを ノイズとみなすか、という尸け持ては坷沸嘿甩にみられる澄唯鼎慷 附据の眷圭、呵介からあるわけではありませんから。慨规が墓袋陵簇を 积って、件侨眶菇陇がある镍刨弓掠拌になると、そういうことが 弹こり评るのではないでしょうか。
もう办つは、票じような弓掠拌のノイズ屯喇尸が2つのニュ〖ロンに 掐った眷圭には、毋えば呵夺の毋でいうと、FitzHugh-Nagumoモデル にそういった弓掠拌の慨规を掐れると、ニュ〖ロンの极券券残の 极甘陵簇が润撅に光くなって、件袋弄になってくるという甫垫が あります[14]。そうすると、このような觉斗を雇えただけで、 ある罢蹋で佰なる坷沸嘿甩の票袋附据のようなものが、ノイズによって 裁庐されるというようなことも弹こりうるのではないかと雇えられる わけです。こういうものは、もちろん海は1つとか2つくらいのニュ〖ロン で雇えているわけですが、それがどんどんつながったときに办挛部が 弹こるかというのは、ちょっとやってみるとおもしろいかなあという ふうに炊じております。
笆惧をまとめますと、まず栏挛にはゆらぎがありまして、これは坷沸废 の替磊り房の瓤炳ˇ瓤纪にはちょっと稍旁圭なのではないかと蛔われる かもしれませんが、涩ずしもそうではない婶尸があって、それは 澄唯鼎慷という矢坍で胺われてきました。そして、その鼎慷の拉墙を 光めるのに墓袋陵簇を积った1/fゆらぎというのは润撅に旁圭がよい ということです。
企つ誊は、これはお箕粗があれば塑碰はお厦ししたかったのですが、 悸はそのメカニズムについても警し慌祸が渴んでいまして、坷沸のダ イナミクスが脚妥であるということです。しかも、ただダイナミックで あるということだけではなくて、いわゆるリラクゼ〖ションの册镍が 侍にあるということ、坷沸栏妄池のことばでいえば陵滦稍炳とか、 そういう雇え数が润撅に脚妥であるということです。
呵稿は、黎ほど拷し惧げたように、蔷ˇ坷沸废における攫鼠コ〖ディング を雇える惧で、ノイズ、あるいはノイズ屯の罢蹋のある慨规の蔡たす 舔充というのは、警し雇えてみたらおもしろいのではないかということです。
笆惧です。
玻怀 ありがとうございます。笆惧の爬につきまして、ご剂啼霹が ございましたら∧∧。
藩付 澎络の藩付ですけれども、叉」やっぱり巫静の惟眷からいう と、黎ほどのニュ〖ラルネットの厦もそうなんですが、ニュ〖ロンの アナロジ〖として、それをモデル步して、ニュ〖ロンと润撅に击た瓢 轮が叫るというのをもう办刨やっぱり∧∧。黎ほどの捍疲黎栏の剂啼 とも簇息するんですが、悸狠にニュ〖ロンのファンクションをどう棱 汤するか、部を瓤鼻しているのかというのは润撅に督蹋を积つんです けれども。黎栏の海のFitzHugh-Nagumoのモデルを蝗われた澄 唯鼎慷のああいったので、毋えば蔷でのニュ〖ラルトランスミッタ〖 による帕茫のどういうような娄烫を瓤鼻しているのか。あるいは肌の 辉摧黎栏のお厦みたいに、ある陕弄な觉轮では1/fのゆらぎが 恃わってくるというようなことと、部か陕弄な觉轮がそこから苞き弹 こされてくるというような、栏妄弄池弄な娄烫と部かつながりがある かどうかということについて兜えていただきたいんですけれども。
怀塑 恫らく企つあると蛔うんですが、办つは、毋えば FitzHugh-NagumoとかHodgkin-Huxleyを蝗うのが、悸狠のニュ〖ロン のモデルになるか、あるいはニュ〖ラルネットは蔷のネットワ〖クの モデルになるかという黎ほどからの厦玛に簇してですが、この澄唯 鼎慷という附据に簇しては、悸狠の坷沸で票じことが弹こるというのが 澄かめられていますし、かなりいい悸赋废が寥まれているのも澄かなん ですね。そこで脱いられたのは球咖花不であります。そして、悸狠 の坷沸のモデルとなるシステムで、いわゆる铜咖花不が铜网に漂く という冯蔡が、海泣の厦のように、毋えばわかったとします。わかった というか、それを悸狠の坷沸で澄かめる涟に、簿の悸赋みたいなものを乖っ たわけです。これを减けて、附哼填宏さんの鼎票甫垫荚は、悸狠の 坷沸を脱いた悸赋废において、海刨は铜咖花不を掐れみて、塑碰か どうか澄かめようというようなことを纷茶しているそうです。で すから、そのモデル步极挛は、警なくともそういう糠しい悸赋を栏み 叫すことには舔に惟つというふうに雇えられるのではないでしょうか。
もう办つは、恫らく、铜咖花不がなぜ叫るかということ、あるい はその墓袋陵簇の刨圭いがどういうふうに陕轮で恃わっていったりとか、 あるいはネットワ〖クの觉轮で恃わっていったりとかすることと簇息 していると蛔うのですが、これはちょっとよくわかりません。という よりも、その弹富は、毋えば铜咖花不に簇しては羔涟面もフラクタルの 厦にもありましたように、栏挛のみでなく湿妄システム链忍にも赂哼 するわけですから、そもそも鼎奶な弹富とかそういうものを滇める ことが澄かに岂しいことなのです。ですから、毋えば桔茨拇泪なら桔茨 拇泪废に、あるいはほかの坷沸拇泪废なら坷沸拇泪废というように、 改侍に玫っていかなければいけないということになるかと蛔うのですが、 そういうような活みは途りよく乖われていないということでしょうか。
玻怀 ほかにございますでしょうか。
底瘦 谨灰板络妈企陕薄の底瘦ですけれども、f尸の1のノイズが 掐ることで润撅に网爬というのを黎栏は动拇されたんですが、嫡に毋 えばお钳大りなんかになりますと、看秋恃瓢なんかでもf极捐に夺い ようなノイズが叫ますけれども、そういった网爬というのは黎栏数と してはやられてはないんですか。というのは、黎ほどノイズがどうい う怠进で叫るかということに簇してはまだいろいろ柒婶で叫る怠进は 岂しいということでしたけれども、裁勿とともにそういうスペクトル が恃わってくるということは恫らくそれぞれ网爬があって、ただ、あ る数烫から斧た眷圭にはf尸の1がf极捐に夺づくということが稍网 に饭いてきて、毋えば祭陕とかに簇息しているのではないかというふ うに讳は雇えているんですが。
怀塑 怠墙弄网爬という烫に簇してはおっしゃるとおりですが、 悸は、ブラウン笨瓢そのものでFitzHugh-Nagumoをドライブしても、 あまり券残しません。もちろんするところでは卖怀券残するんです けれども、しないところでは链くしないので、链挛として斧ると 掐叫蜗の陵簇はないんです。ただ、ブラウン笨瓢极挛は裁恕册镍として 润撅に络きな墓袋陵簇を积っていますよね。ですから、陵簇だけが 脚妥なわけではなくて、ある镍刨陵簇を积っていて、そしてある镍刨、 恃な咐い数ですけれども、ギザギザしている、その面粗ぐらいが 攫鼠のコ〖ディング墙を笼すという爬からはほどよいということだと 蛔います。そして、そのほどよいノイズが栏挛に首哼する ということ极挛がなぜかという爬に簇しては、润撅に督蹋考い啼玛 であろうと蛔います。毋えば裁勿でノイズ屯喇尸の陵簇がブレ〖ク ダウンしていってブラウン笨瓢みたいになっていくと、海みたいな 跟蔡が簿に铜跟に侯脱しているとすれば、それは禄なわれる材墙拉が あるわけです。悸狠毋えば、夺钳啼玛となっている啪泡の啼玛と 簇息して、光勿荚の颅婶炊承とかそういうものを、盖铜炊承达にノイズを 裁えることによって光めてやろうというような炳脱が幌まっている とも使いております。まだうまくいっているかどうかわからない ですけれども。
玻怀 よろしいでしょうか。
ありがとうございます。
(≈给倡シンポジウム ∝ホメオスタシスの附哼(いま)≠. 士喇9钳刨彩甫锐 答茸Bˇ措茶拇汉 ∝桔茨废のホメオスタシスとその佰撅の答塑付妄に簇する 糠しい琵圭弄アプロ〖チ≠(甫垫洛山荚, 络柒坝盗: 澎络板池婶戏钳陕彩)による, 澎叠, 1998钳2奉.∽より)
3≥4 栏挛ゆらぎと坷沸废のダイナミクス
This document was generated using the LaTeX2HTML translator Version 96.1 (Feb 5, 1996) Copyright © 1993, 1994, 1995, 1996, Nikos Drakos, Computer Based Learning Unit, University of Leeds.
The command line arguments were:
latex2html -split 0 jres_9.
The translation was initiated by Yoshiharu Yamamoto on 1998钳06奉06泣 (炮) 14箕25尸24擅 JST